


Handbook of Nascent Human Beings: A Visual Aid for Understanding the Science and Experimentation


This is Issue 8 of the On Science series.
Originally published February 25, 2022, and has been updated on July 7, 2022 to include companion Table 1.
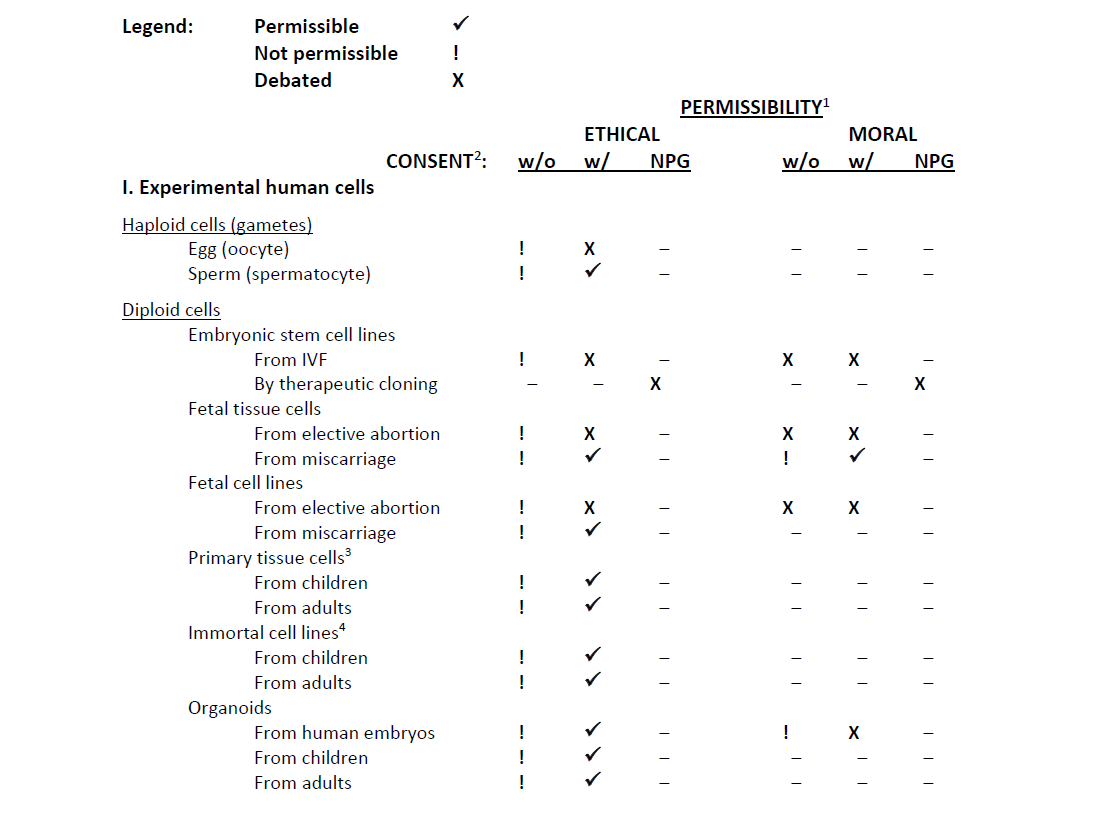
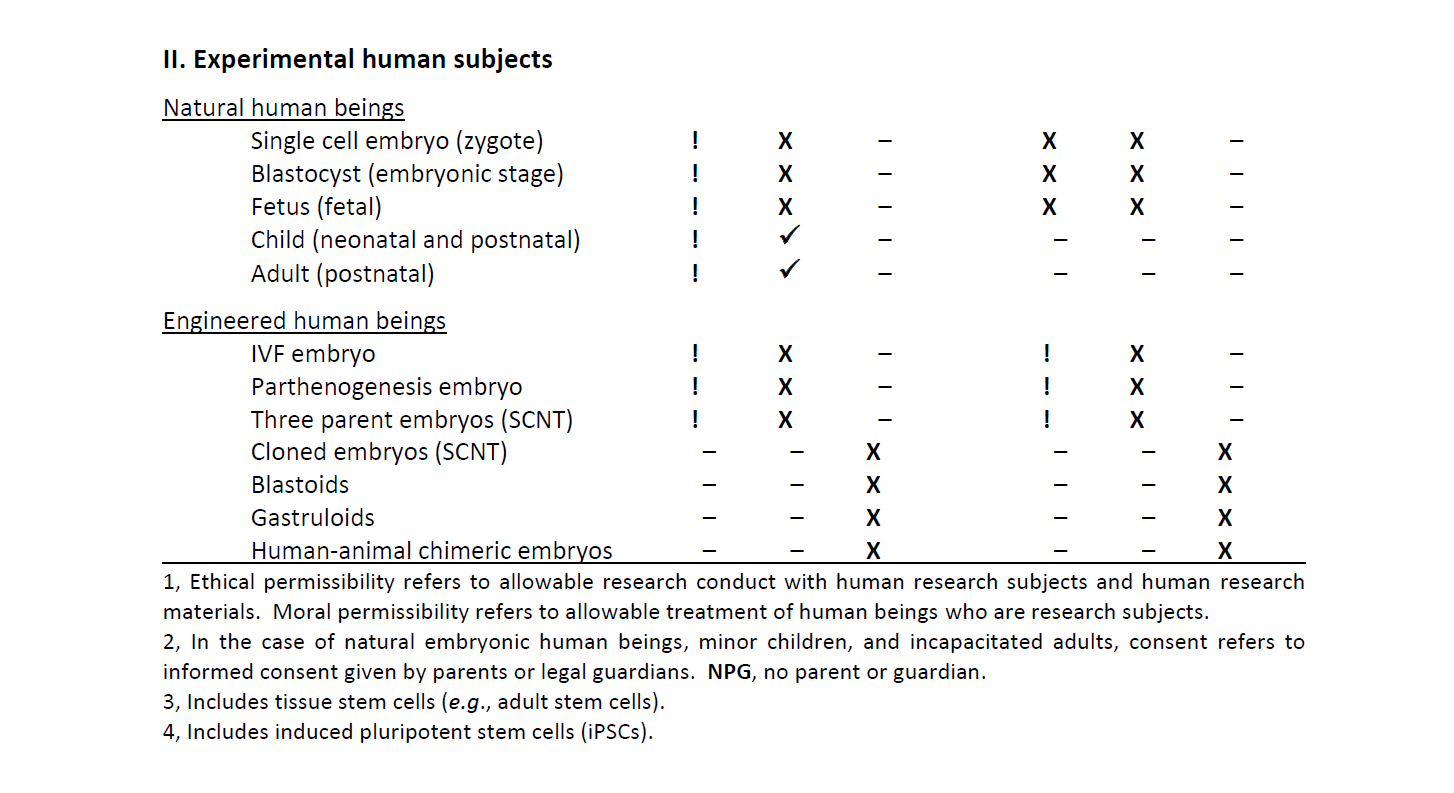
To view Table 1 below as a PDF, see: Table 1. State of the Ethical and Moral Permissibility of Human Experimentation Research
Introduction to the Handbook
The purpose of this handbook is to provide a useful reference guide to understanding the biological nature and properties of nascent human beings and cells derived from nascent human beings.
The Handbook is organized by the stages of human development advancing from conception toward adulthood. At each stage of development, human beings and their cells conceived by natural processes are compared to those conceived by artificial means involving bioengineering, modification, and/or experimental manipulation. Living organisms and their cells that are produced by any type of “artificial” means does not necessarily connote that they are unnatural. Their origin and scientific inspiration are still from human organisms and/or biological functions occurring naturally in nature. Scientists have discovered diverse methods for producing both nascent human beings and cells derived from nascent human beings with the aim of scientific experimentation. Any nascent human being produced using artificial means, and the cells derived from them, have the essential nature and properties of human beings and their cells conceived by natural processes.
Human beings conceived by scientific bioengineering are no less human than those conceived by natural processes. Therefore, they have the same moral significance; and they require the same bioethics considerations before use for biomedical experimentation.
Nascent Human Beings Conceived by Natural Processes
Ovulation and Fertilization
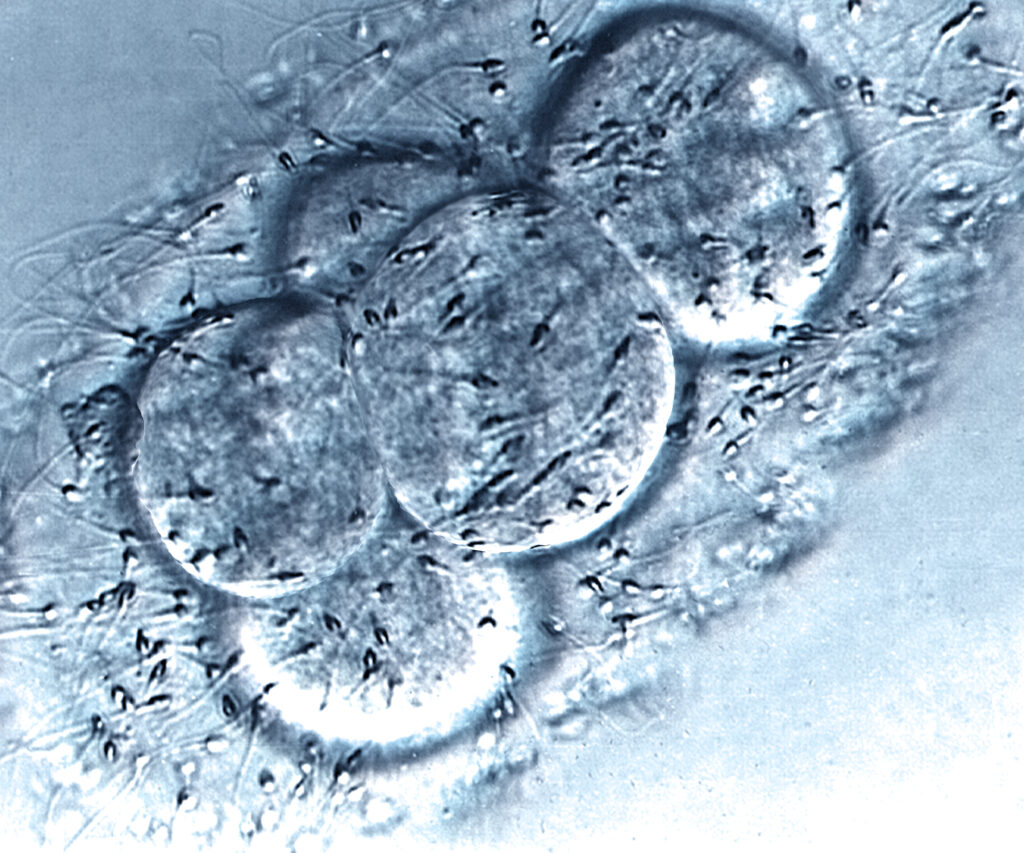
During week 1 (days 0 to 6), an egg (oocyte) and sperm fuse effectively, and a nascent human being is conceived by a process called fertilization. The single-celled nascent human being is called a human zygote. The human zygote is also called a fertilized egg. Zygotes form naturally in the female reproductive tract when a human sperm fertilizes – fuses with – a human egg. This event restores a full-complement of human DNA in the specialized cytoplasm of the egg gamete. This is the crucial event that brings humans into being. Thereafter, given supportive nutrients and conditions, which are ideal in the human uterus, the human development program unfolds spontaneously and autonomously.
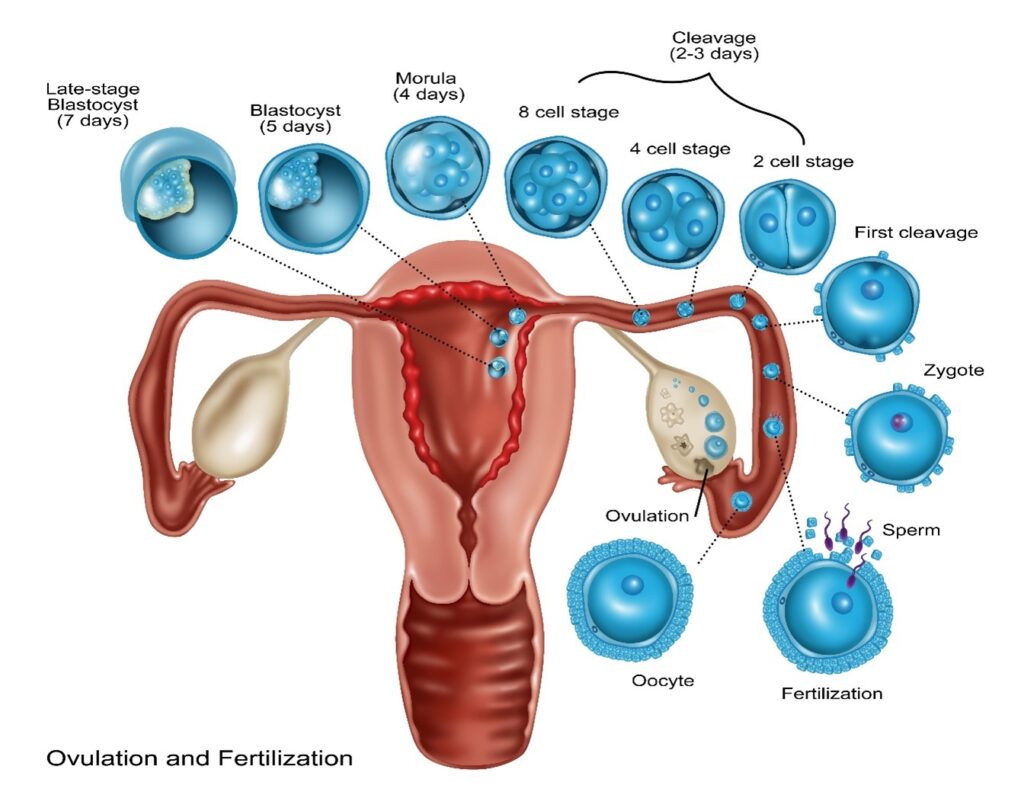
With time and the supportive conditions of the female uterus, natural nascent single-celled zygotic human beings divide into 2 cells, then 4 cells, then 8 cells, and so on until reaching the morula stage. This is the first of trillions of sequential cell divisions in the development program by which they will grow and transform into complex multicellular adult human beings. This stage of development is called embryogenesis, and the nascent human beings of this age are called embryos. Zygotic human beings are single-celled embryonic human beings. The descriptive terms embryo and embryonic are applied until later times in development when human beings achieve the age of their development defined as fetal.
Implantation
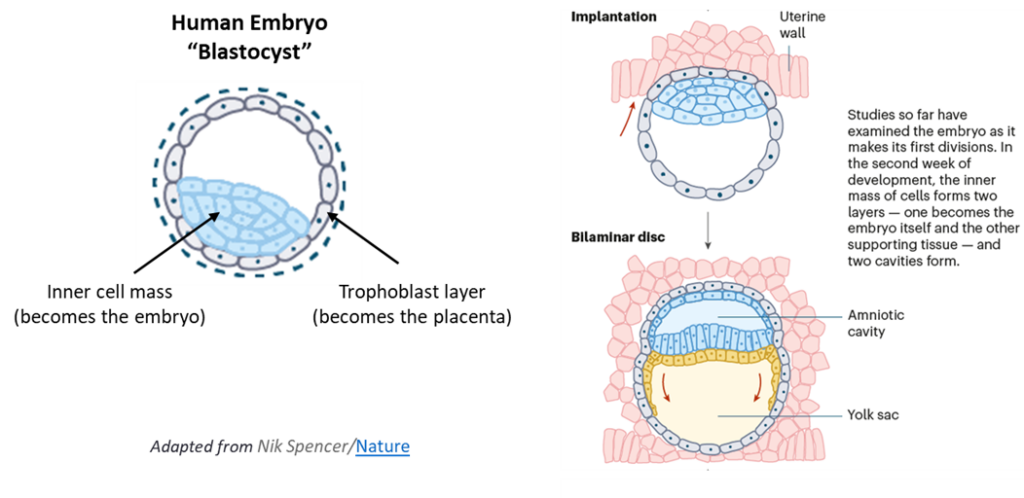
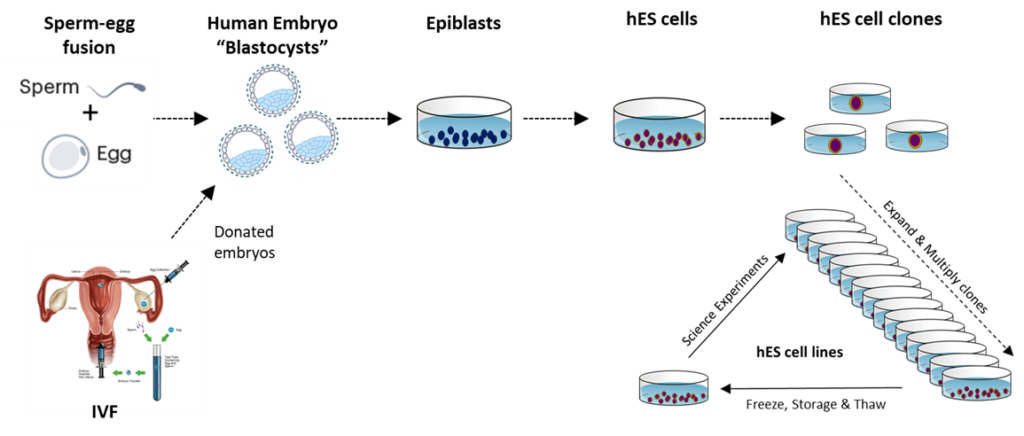
Scientists have defined successive ages of developing embryonic human beings that describe their increasingly complex morphology and functional capabilities. At first is the morula stage, followed by the blastula stage. In their blastula stage of development around day 4-5, nascent human beings are called blastocysts. When of blastocyst age, nascent human beings have the form of a hollow sphere of cells with an inner cell mass that adheres asymmetrically to one side of nascent human beings’ spherical exterior. The cells that compose the inner cell mass are called epiblasts. A late-stage blastocyst then implants into the uterine wall during week 2 (days 7 to 13), which is the beginning of uterine nourishment.
Gastrulation
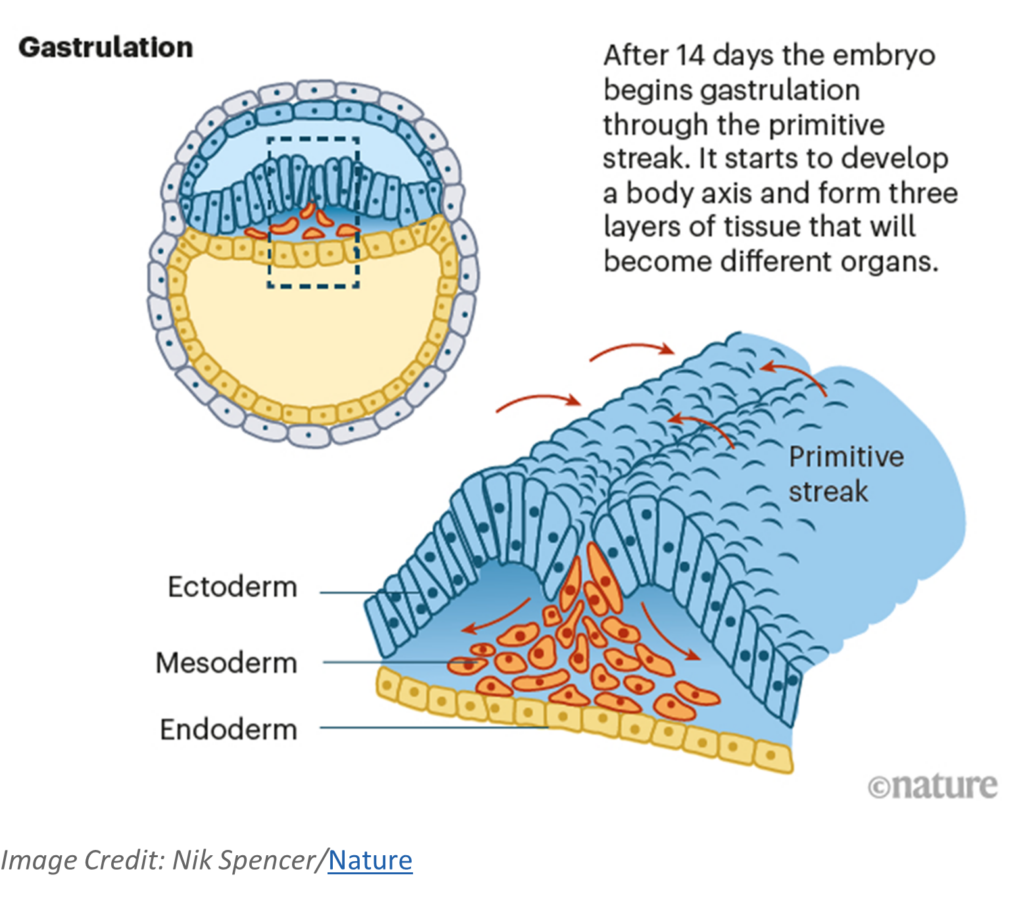
The next age of human development, which has recently received increased attention for human experimentation, is gastrula. Gastrulation is a period of migrations by the cells of blastula age human beings. In calendar age, gastrulation begins during week 3 (days 14 to 20). Its start is signaled by a feature called the primitive streak. The cell migrations originate from the primitive streak to establish three special layers of cells that are the foundation for the development of organs, including the heart, nervous system, and the brain whose development begins soon thereafter. The three layers (ectoderm, mesoderm, endoderm) are called the germ layers. All organs found in human beings when they are of child and adult age developed from one of these layers.
Neural Tube Formation
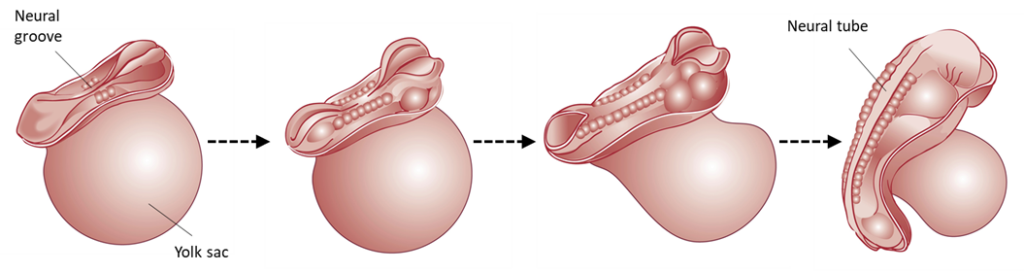
During week 4 (day 21-27) the neural system (brain, spinal cord, nervous system) starts to form from a structure called the neural tube. The neural plate folds to a neural groove that fuses along its length to form the neural tube. Once the neural tube closes, the more specific regions and structures in the brain begin to form and fall into place (forebrain, midbrain, hindbrain). Neural circuits, or connections between neurons that communicate information throughout the brain, are formed in the fetal period. (The brain is fully developed as an adult at around 25 years of age.) The heart starts beating rhythmically during the fourth week as well, around day 22.
The developing nascent human being will undergo dramatic changes the remaining 36 weeks until birth. For more information, please visit our website on prenatal development “The Voyage of Life.”
Cells of Nascent Human Beings
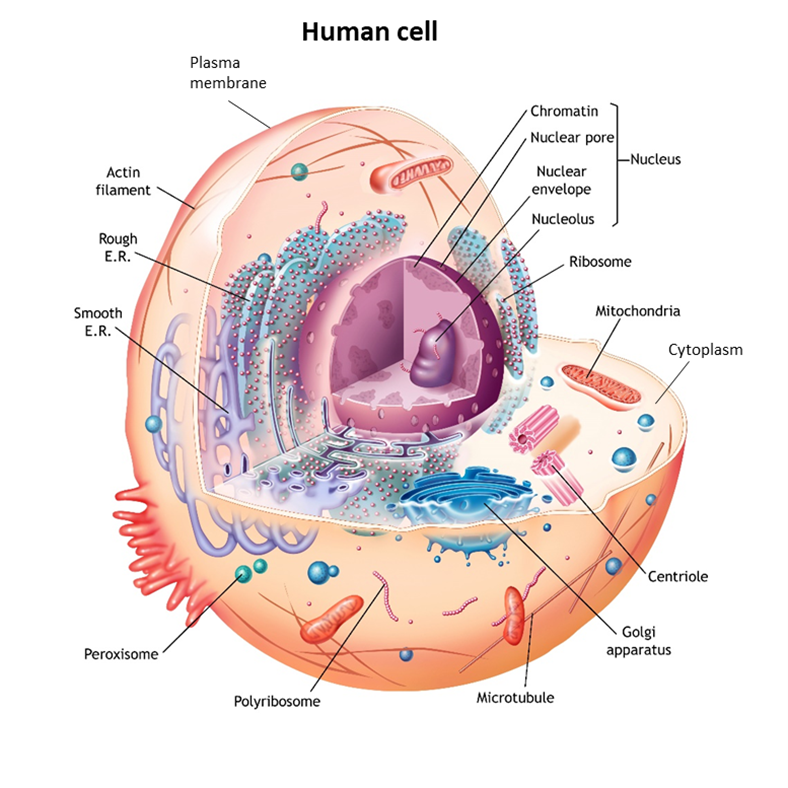
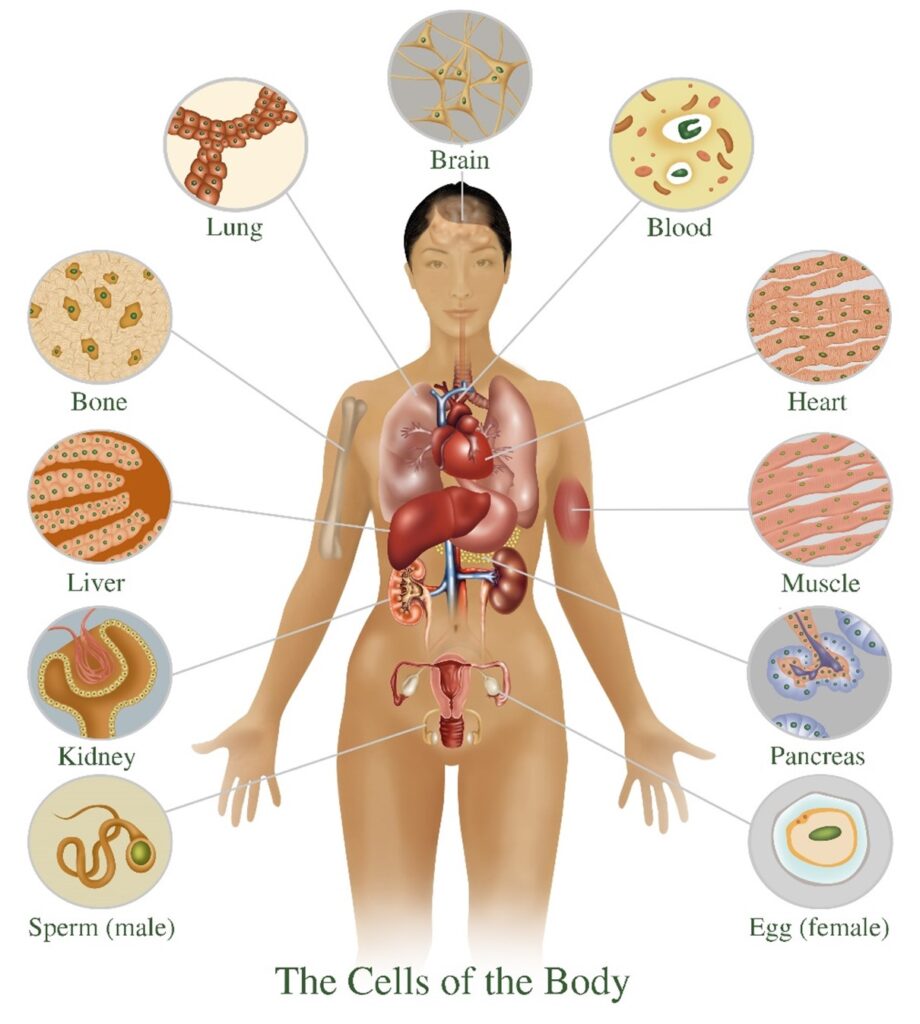
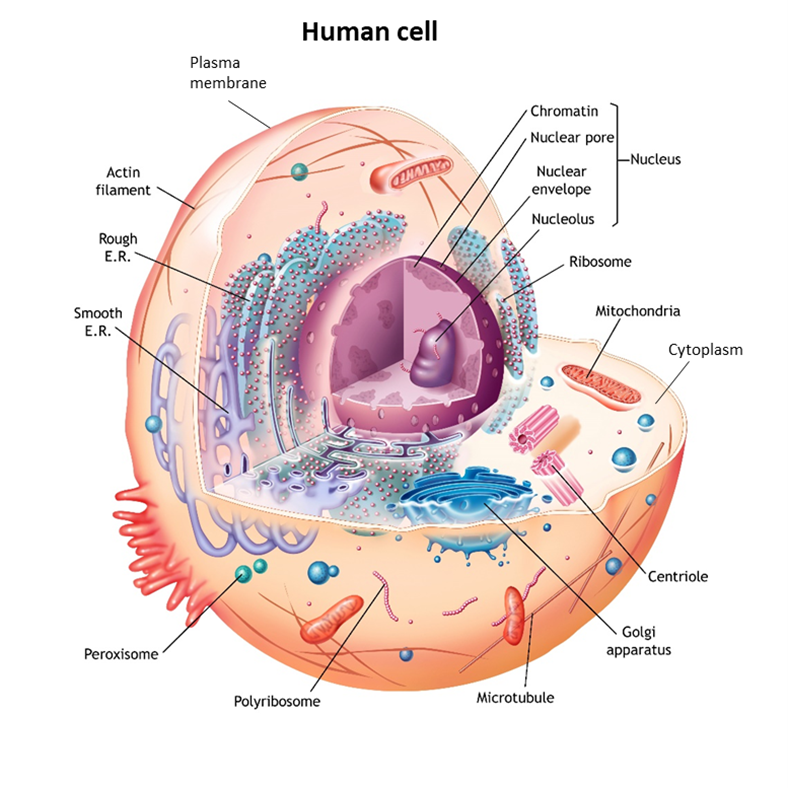
In living organisms, plants and animals, including human beings, cells are the microscopic functional unit of tissues and organs. In nature, there are also many examples of free-living single cells. Examples are amoeba, bacteria, and even yeasts that are used for baking and making wine and beer. The human body at all stages of development and maturation, except at the moment of conception when a nascent human being is only one cell in size, is composed of increasing numbers of cells that perform the many complex functions of human tissues and organs.
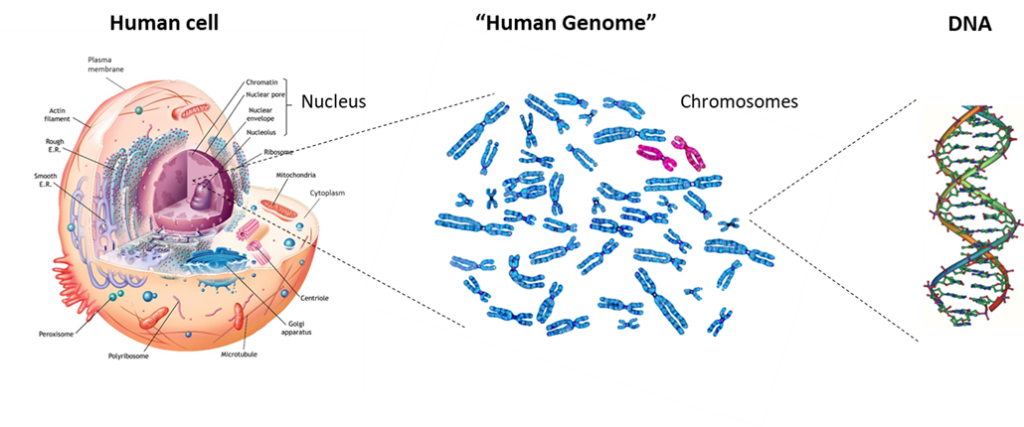
Although there is a vast number of different types of cells in nature and in the human body, cells as units of life share the same fundamental features. They have a plasma membrane that is like a double-walled balloon that is filled with their cellular contents. Their gelatin-like contents generally occur in two distinct compartments. Typically, the larger compartment is the cytoplasm (“the cell stuff”). The second, smaller compartment is suspended within the gel of the cytoplasm. Called the nucleus, the second compartment contains the cell’s genetic material, the DNA tightly packaged in the form of chromosomes. Although there are myriad differences in the detailed aspects and properties of these three basic cellular features from cell to cell, they are fundamental to living cells, including those of human beings.
The basis for determining whether a cell is alive is different than the basis for determining whether a mature human being is alive. The basis for determining whether a mature human being, with developed tissues and organs, is alive is by medical determinants. The criteria include three attributes: heartbeat, breathing, and brain function. These medical criteria establish whether a human being with developed tissues and organs can continue to function in a manner that is biologically sustainable either autonomously or through medical intervention.
Biological sustainability is the ability to keep the many different cell types of the body alive. When human beings are at a stage of development for which medical criteria can be applied, their medical death is rapidly followed by the essential cells of their bodies dying as well. However, if provided essential nutrients and conditions, the cells of a medically deceased human being can be kept alive for considerable periods of time using modified culture techniques. This property of human cells makes organ transplants, stem cell transplantation medicine, and human cell research possible.
At earlier stages of human development before medical criteria for being alive are applicable, biological sustainability is the basis for evaluating whether human beings are alive. Biological sustainability is the same criteria applied to determine whether any individual cell or organized groups of cells are alive. When cells are alive, they are able to take up energy from their environment and convert that energy into cell work. Sources of energy can be solar or chemical. Plant cells have the ability to absorb and use solar energy. Animal cells, including human cells, get chemical energy by breaking the chemical bonds of the nutrient compounds they absorb from their environment. They convert the chemical energy into many different forms of cell work that are specific to their function. Types of cell work include functions like breaking down compounds; making body components and products like hair, nails, and sweat; generating nerve impulses; and expressing the genes in their DNA.
An essential type of work that is a quintessential property of many types of living cells is the ability to duplicate their DNA (called DNA replication) and divide to make two new cells. This cell division is crucial for human beings to grow from their single-celled stage of development to their mature fully developed adult body stage. The cellular process by which the many trillions of cells produced by division transform into cell types with many varied functions is called differentiation. Cell divisions continue throughout the tissues and organs of fully mature human beings. Differentiated functional cells in fully developed adult human beings lose the ability to divide. They get old and die, or sometimes they are just lost from the body by natural processes of physical wear. Continuous cell division by specialized cells in the body serves to replace dead and lost differentiated cells.
Experimentation Using Cells from Nascent Human Beings
Human Cell Lines
Both natural and bioengineered human-derived cells have been developed for biomedical research. Examples of natural human-derived cells include tissue and organ cells that are removed from human beings for either direct experimentation or for propagation by supporting their cell division in specialized cell cultures used for later experimentation. Cell culture propagation enables more expansive and longer-term biomedical investigations for days to months.
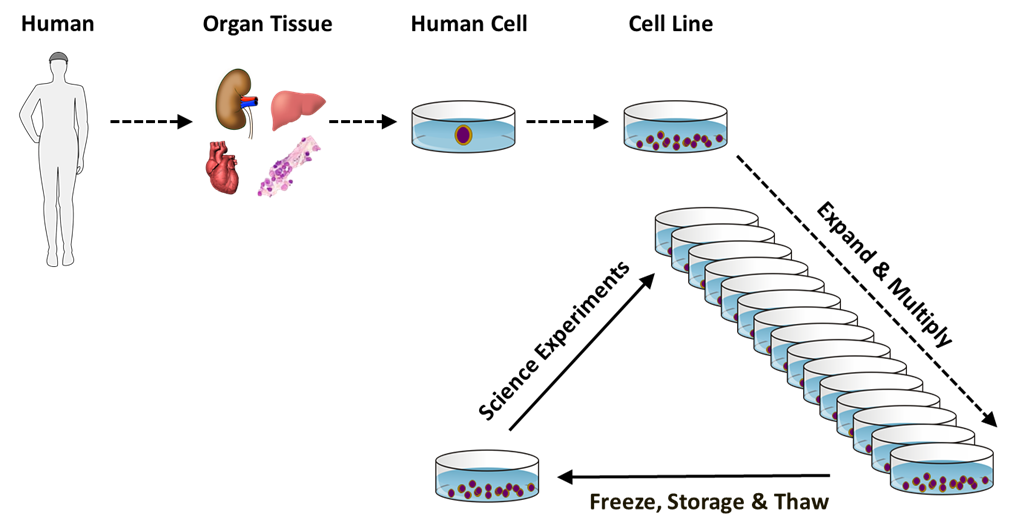
Natural human cells typically lose their ability to divide after only a few months of culturing. Because of this characteristic experimental limitation, natural human cell cultures are sometimes called mortal cell strains, referring to the inevitable “death” of the cells with extended culturing. Despite their culturing limit, natural human cells can be stored for use over many years by freezing the cells under conditions that allow thawing and their subsequent return to limited culturing.
Natural cell cultures have been developed from both normal human tissues and diseased human tissues, like tumors. Tumor cell cultures have been of particular interest for cancer research and because many of them can be cultured more extensively without the division limit of mortal cell strains. For this reason, many human tumor-derived cell cultures are described as immortal cell lines. One well known example is the HeLa cell line.
Generally, bioengineered human-derived cells have the extensive culturing capacity of immortal cell lines, though they may or may not have other properties of tumor cells. Such cells are bioengineered, often genetically, to provide new properties of interest or utility for scientific investigations. One of the often-engineered properties is the ability to propagate in culture as immortal cell lines.
Another important attribute of natural and bioengineered immortal cell lines is their clonality. All mortal cell strains are composed of cells that are the descendants of the divisions of multiple cells in initially isolated populations of human tissue cells. Because cellular heterogeneity can undermine the quality and interpretation of biomedical experiments, scientists prefer to conduct experiments with cell cultures that were produced from lineages of cell divisions that began with a single isolated cell. Human cell cultures developed from a single originating cell are referred to as clonal cell lines. Because of their limited division capacity, cells from cultures of natural mortal cell strains are unable to produce either clonal mortal cell strains or clonal immortal cell lines. In contrast, cells from cultures of many different types of natural immortal tumor cell lines and bioengineered immortal cell lines can be, and have been, used to produce respective clonal immortal cell lines. With modern technologies, bioengineered immortal cell lines can be developed with the cells of fetal, child, or adult human beings, whether living or deceased, from virtually any organ or tissue.
For more information on how cell lines (human and non-human) are used in vaccine development and production, please refer to our FACT SHEET.
Human Gametes (Sperm and Egg)
Human gametes are natural specialized cells that fuse together to form a human being. The fundamental distinction of gamete cells from all other cells of human development and in the mature human body is that their nucleus contains only half the amount of DNA found in other cells. By a specialized cell division process called meiosis, which occurs in the ovaries and in the testes, some cells divide without first replicating their DNA. As a result of this reductive cell division, their DNA content is reduced to one half of the usual complement in the cells of human beings. In the female ovary, gamete cells are called oocytes or more generically, eggs. In this half-complement DNA state, they are also called unfertilized eggs. In the male testes, gamete cells are called spermatocytes, or more generically, sperm.
Like other human cell types, for use in scientific experimentation and biomedical research, natural gamete cells are obtained from human donors; and though there are some exceptional situations, donors are generally of legal age. Because sperm are produced throughout the life span of men after they reach reproductive age and do not require medical procedures for collection, human sperm donation is a common practice. Donation of natural human eggs requires that women who have achieved reproductive age undergo hormonal treatments and medical procedures that involve significant risks of pain and injury.

Bioengineered human gamete cells have recently been developed (See page 24, “In Vitro Gametogenesis”). Specialized types of cultured human cell lines were bioengineered to undergo meiosis with reductive cell divisions to produce human sperm and eggs under experimental conditions.
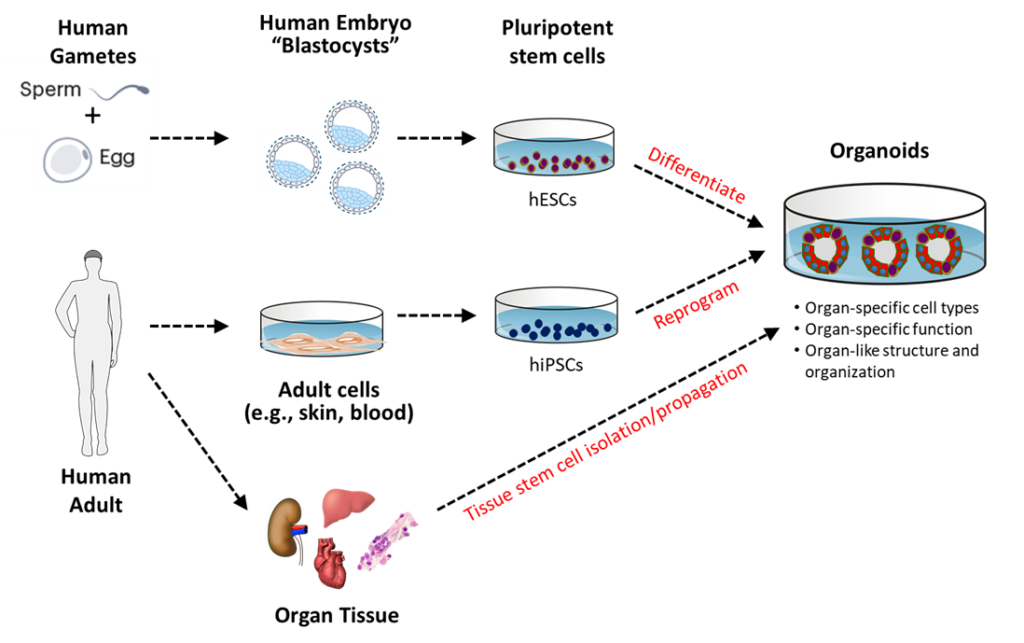
Embryonic Stem Cells and Embryonic Stem Cell Lines
Human embryonic stem cells (hESCs) are derived in culture from the epiblast cells of embryonic nascent human beings after they complete their blastocyst development. The culture conditions used prevent the epiblast cells from continuing their human development while still maintaining their high proliferative capacity. Embryonic stem cells can be produced in large numbers and used to make clonal hESC strains. Human embryonic stem cell lines are available for NIH-funded research. For more information, see the NIH Human Embryonic Stem Cell Registry.
Induced Pluripotent Stem Cells
Another scientific development initially had a major impact on reducing the production of hESCs. In 2006, Dr. Shinya Yamanaka reported the first production of induced pluripotent stem cells (iPSCs). Yamanaka led his research team in the discovery of sets of genes that could be introduced into mature somatic mouse cells and induce them to regain properties nearly identical to those of mouse ESCs. It was not very long, thereafter, that the labs of Yamanaka and others showed that the same types of genes could be used to make human iPSCs (hiPSCs) by similar techniques, using mature somatic human cells (e.g., skin cells). In 2012, Yamanaka was awarded the Nobel Prize in Physiology or Medicine for this scientific advance.
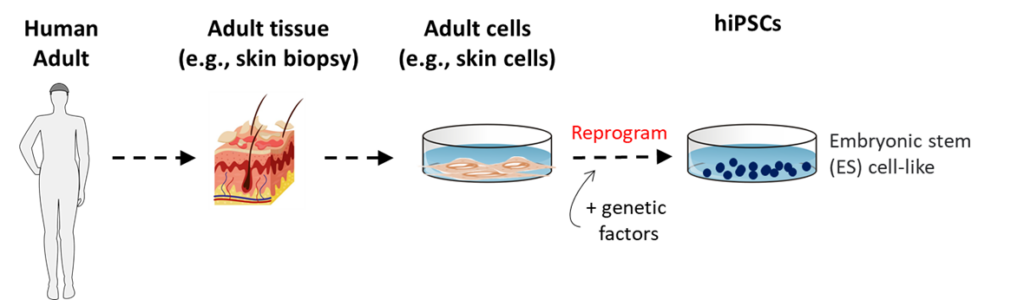
In the early days of iPSC experimentation, scientists performed many analyses to look for important differences between iPSCs and ESCs. No significant differences have been reported. For a period, and continuing to a small extent currently, experimentation with hiPSCs included a smaller set of experiments with hESCs for comparison purposes. However, with time, for a while most studies with hiPSCs had progressed to primarily use of hiPSCs alone. Now, more recently in reports of bioengineered, cloned embryonic nascent human beings, both hESCs and hiPSCs have been involved (See page 30, Human Embryo Models).
A large focus of hiPSC experimentation is finding ways to control and use the remarkable developmental capability of pluripotent cells to develop innovative medical therapies for organ and tissue replacement. Examples of this research include scientists using hiPSCs in attempts to make mature insulin-producing pancreatic cells that cure child human beings and adult human beings of Type I diabetes; laboratory tissue engineering to produce organs and tissues for transplantation medicine; and bioengineering the development of human organs for transplantation in chimeric animals like pigs (See page 34, Human-Animal Chimeric Embryo Research).
Because hiPSCs are biologically equivalent to hESCs except for their cells of origin, attempts to develop medical therapies with hiPSCs have run into the same difficulties posed by hESCs. Human iPSCs also form teratomas when they are introduced into mature organs and tissues of the body; and when directed to develop in laboratory experiments, they also preferentially produce under-developed cells with fetal organ and tissue properties, lacking the child and adult properties that are needed for organ and tissue transplantation medicine.
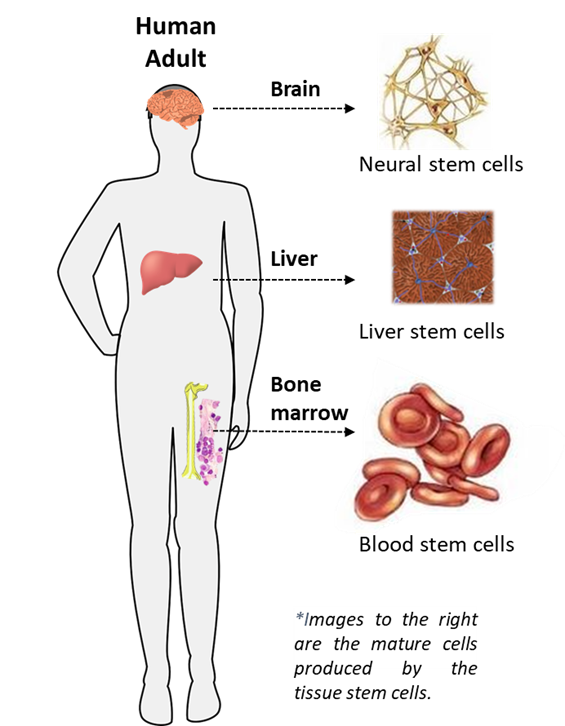
Tissue Stem Cells
Although organs and tissues in child human beings and adult human beings appear static, they are turning over constantly. In nearly all organs and tissues, the oldest, and functionally most mature cells are no longer able to divide. With lifetimes of weeks to months, they die and are removed or lost. Their continuing loss is replenished by the division of younger cells. The immature cells that produce these newer generations of organ and tissue cells are tissue stem cells.
Tissue stem cells first develop in human beings during their late fetal development as distinctive organs and tissue begin to form and mature, and then they are maintained through the development and maturation of child human beings and adult human beings. Tissue stem cells continue to function in the organs and tissues of aged human beings even after human beings’ medically certified death. For this reason, organs and tissues of deceased human beings can continue to function normally after transplantation into a living recipient.
Tissue stem cells have a number of other names, including for example adult stem cells, somatic stem cells, and tissue-specific stem cells. They have distinct biological properties from hESCs. First, unlike hESCs, they are naturally occurring and have not been bioengineered or experimentally
derived. They are normal human tissue cells. They are not pluripotent, meaning they are able to produce only the distinctive cells of the organ or tissue that they renew. If the organ or tissue they renew has only one mature cell type – for example the lens cells of the eye –, they are termed unipotent tissue stem cells, because they produce only one cell type that is more maturely developed, or “differentiated,” than themselves. If the organ or tissue renewed has multiple types of mature cells, – for example the blood system that has many different types of mature functional cells – they are termed multipotent tissue stem cells, because they produce more than one cell type that is more differentiated than themselves. Because tissue stem cells are always much less functionally differentiated than the mature cells they divide to produce, they are described as primitive compared to their differentiated progeny cells.
The most important biological difference between tissue stem cells and hESCs is the manner in which they produce mature differentiated tissue cells. Tissue stem cells renew tissues by a process called asymmetric self-renewal division. Their divisions are asymmetric, producing one new cell that is a new tissue stem cell; and another new cell that either differentiates directly or divides to produce other differentiated cells that form the organ or tissue but eventually stop dividing permanently. In this way, tissue stem cells are able to renew differentiated tissue cells in mature organs and tissues continuously without losing their crucial stem cell identity and function. During asymmetric self-renewal divisions, tissue stem cells also have a special form of DNA called immortal DNA strands that protects their DNA genome from replication errors (i.e., gene mutations). The asymmetric self-renewal organization of human tissues results in tissue stem cells being a small fraction compared to the many cells they continually produce. Their fraction in human tissues ranges from 1 in several thousand cells to as little as 1 in one hundred thousand cells.

Like tissue stem cells, hESCs are defined as stem cells because of their ability to both renew themselves (“self-renew”) and differentiate. However, unlike tissue stem cells, hESCs cannot, and do not, achieve these two essential criteria simultaneously by asymmetric self-renewal division. When hESCs self-renew, they produce more hESCs by symmetric self-renewal divisions. The symmetric self-renewal divisions by hESCs produce two new hESCs. The experimental conditions that allow hESC self-renewal prevent their formation of differentiated cells. When hESCs are switched to conditions that allow their differentiation, hESCs do not asymmetrically produce other cells that differentiate. Instead, they differentiate directly themselves. This intrinsic biological property of hESCs is generally understated by scientists who promote their potential for biomedical research. Yet, it precludes them from ever being used as therapeutic cells for use in organ and tissue transplantation medicine. Human ESCs could never work to renew human tissue cells like tissue stem cells do naturally.
Tissue stem cells have been the focus of biological investigations of their nature and function in human organs and tissue for more than half a century. A major focus has been elucidating their role in human carcinogenesis when they sustain either spontaneous or induced mutations in cancer-causing genes. Germane to this Handbook’s purpose, even before iPSC technology, research with tissue stem cells was an important counterbalance to the claims of the benefits of hESC research.
Tissue stem cells are a viable alternative to hESCs for developing innovative cellular therapies for diseases and disorders that lack effective medical treatments. Biomedical research with tissue stem cells continues to focus on their well-known challenges, which like hESCs reflect their distinctive biological origins. Because they are a rarified population of natural tissue stem cells, unlike hESCs, it has been difficult to study them in large and purified numbers. Their intrinsic asymmetric self-renewal division makes it difficult to produce them in large quantities for research investigations, tissue engineering of organs, or transplantation medicine. Efforts to overcome this biological limitation of tissue stem cells are underway in many academic and industrial laboratories.
Despite the current challenges in tissue stem cell medicine, this alternative field is well beyond hESC research in medical practice. Unlike hESC experimentation, which has yet to yield proof of concept for cellular therapeutics, human tissue stem cell experimentation established proof of its therapeutic concept with bone marrow blood (hematopoietic) stem cell and umbilical cord blood stem cell transplantation treatments decades ago. Current human tissue stem cell medical research is building on this foundation.
Metakaryote Stem Cells
The most recently defined stem cells are metakaryote stem cells. They appear during fetal development and are responsible for the initiation of mature tissues and organs. They have many of the special properties of tissue stem cells, and may even be the developmental precursors of tissue stem cells. However, they have several unique properties. Like tissue stem cells, they produce fetal organs by asymmetric cell divisions. However, their cell division proceeds without the usual breakdown of the nucleus and alignment of the duplicated chromosomes for separation. Instead, metakaryote stem cells have characteristic “bell-shaped nuclei” that divide directly without first opening to visibly align the duplicated chromosomes.
Metakaryote stem cells are very rarely seen in the normal organs and tissues of child human beings and adult human beings. However, they do appear at high frequencies in tumorous tissues and in the wounds of injured organs and tissues. The significance of their presence in these abnormal states is not fully understood; but in the case of cancers, metakaryote stem cells may be a type of cancer stem cell. Cancer stem cells are responsible for renewing tumor cells much in the same way that tissue stem cells renew normal adult tissue cells, and normal metakaryote stem cells renew normal fetal tissue cells.
The field for human metakaryote stem cell experimentation is still in its infancy. The first isolations of the cells from human tumor tissue only occurred in the recent decade. Among their many important biological properties, they are the only human tissue stem cells that are specifically identifiable in tissues or in the laboratory. The bell-shaped nuclei of their metakaryotic cell divisions are morphologically distinctive, making it possible to identify them in organs and tissues and to quantify them in experiments. They are the only type of tissue stem cells with this capability. Current research is focused on defining their potential as advantageous targets for cancer treatment and accelerated wound repair.
Organoids
Scientists use the term “organoid”, meaning organ-like, to indicate when the experimental cellular structures they produce in cell cultures have more features of natural organs than found in traditional tissue cell culture systems. The first organoids were produced from the experimental reassembly of dissociated human tissues. Like mammalian cells in general, human organ and tissue cells have instructions on their surfaces that allow them to reorganize after dissociation if the conditions are conducive. Organoids are not little whole organs. They are microscopic partial organ structures that retain some of the biological and physiological properties of an intact organ. Organoids allow scientists to study organ functions in the laboratory that are inaccessible or difficult to study in intact organs in the intact body.
Many complex functions of the human body that can be important in disease and medicine require organized structures composed of distinctive mature types of cells. A shortcoming of conventional human cell cultures (e.g., immortal cell lines) is the loss of this important higher-order organization. Because organoid culture systems provide this advantage, their use for biomedical research has increased exponentially in the past decade.
Another driver for the recent explosion in human organoid experimentation has been the introduction of the use of hESC lines and iPSC lines to produce the differentiated cells that are assembled to form organoids corresponding to many different human organs of medical importance. Most recently these disturbingly include human brain cerebral cortex organoids and “gastruloids” (See also page 30, Human Embryo Models). Gastruloids are iPSC-produced organoids corresponding to embryonic nascent human beings of gastrula age. The main innovation motivating scientists is their ability to make genetic manipulations in the originating hESCs and iPSCs, which is facilely done by CRISPR gene editing (See later page 32, Genetic Engineering), and then studying their effects in tissues that approximate the functions of the corresponding human organ or embryonic nascent human beings, respectively.
Nascent Human Beings Conceived by Artificial Processes
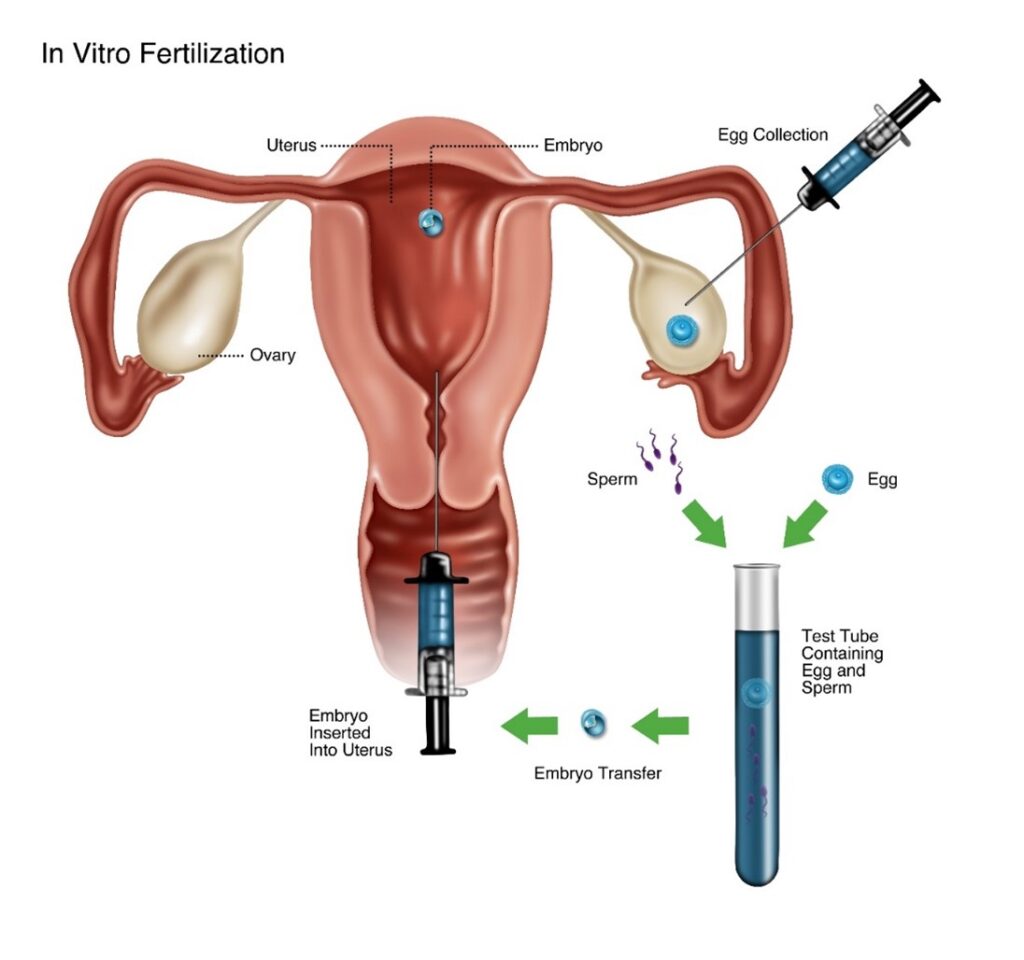
In Vitro Fertilization
As described in Section I, a nascent human being is conceived when an egg and sperm fuse effectively. The creation of single-celled human zygote restores a full-complement of human DNA, a crucial event that brings humans into being. Zygote formation outside the natural environment is the experimental foundation of assisted reproduction by in vitro fertilization (IVF). Zygotes (i.e., fertilized eggs) are produced by combining donor eggs and donor sperm in the lab and allowing them to fuse. Sometimes the sperm is injected directly into the egg to assist the process. This technique is called Intracytoplasmic Sperm Injection (ICSI). The resulting zygotic nascent human beings can be either implanted directly into the uterus for continued development and maturation to birth or frozen for implantation at a later time. Embryo adoption occurs when excess frozen embryonic human beings conceived by IVF are adopted by and implanted into a different woman than their biological mother.
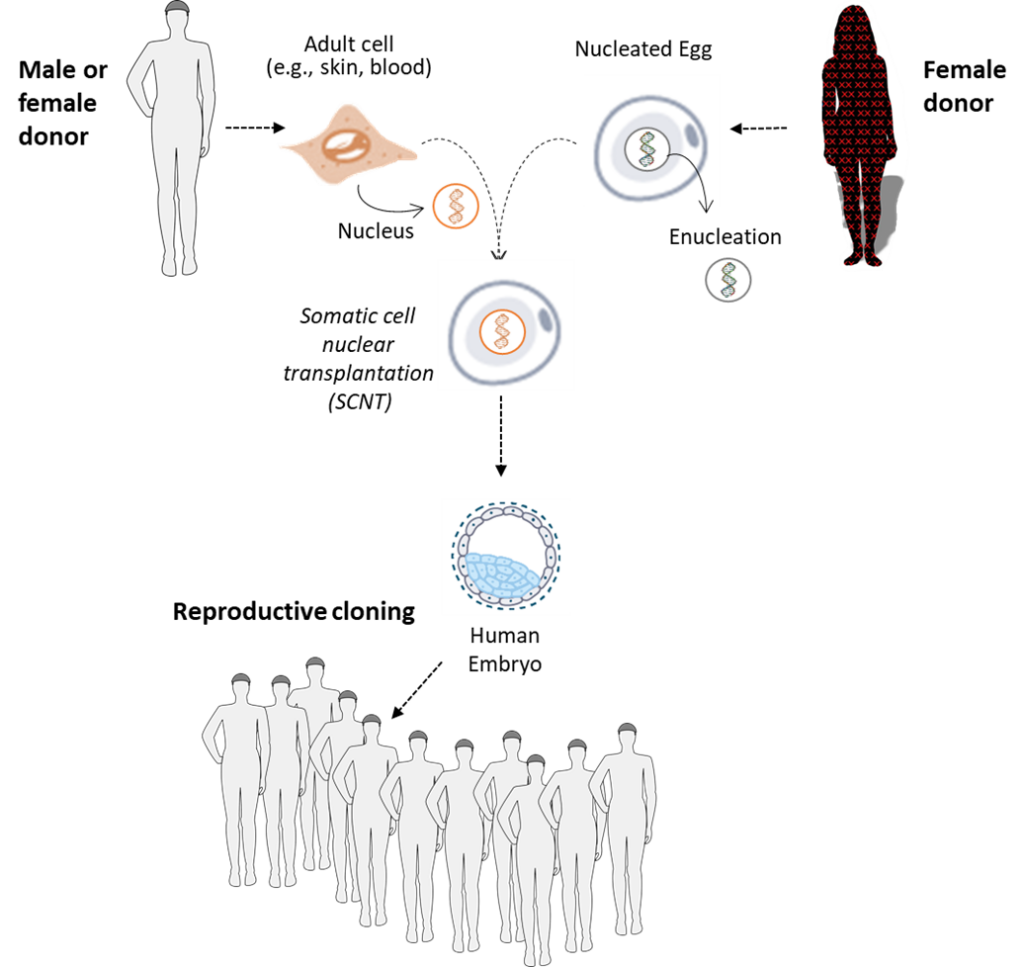
Somatic Cell Nuclear Transfer (Human Cloning)
Scientists have also discovered that whenever a nucleus containing a full-complement of human DNA is introduced into the cytoplasm of an unfertilized human egg, whose nucleus containing a half-complement of DNA was removed beforehand, a nascent human being may be conceived. The nuclei used are isolated from other human body cells or from cultured human cells. This experimentation procedure is called somatic cell nuclear transplantation or SCNT (described and illustrated in more detail below). The term “somatic” categorizes most of the cells of the human body (the “soma”) in contrast to the smaller populations of specialized cells responsible for reproduction. The contrasted cell types are the gamete cells and gamete-producing cells. Collectively they are called the germline cells of the body.
SCNT is known more commonly as human cloning. SCNT is called “cloning” because by fusing multiple eggs for the same woman donor with nuclei from cells of a clonal human cell line, it is possible to conceive zygotic nascent human beings who are genetically identical. Identical because the DNA genetic information in all the inserted nuclei was produced by copying of the DNA in the first single cell that divided to produce the cultured immortal cell line.
Cloning is the procedure by which the now famous “Dolly the Sheep” and her siblings were conceived. Nuclei isolated from cells from clonal cultures of immortal sheep cell lines were inserted into sheep oocytes after removal of the oocytes’ nucleus containing a half-complement of sheep DNA. “Clonal lambs” were born after implantation of the cloned sheep zygotes into the uterus of Dolly’s mother.
Reproductive Cloning
Reproductive cloning is performing with zygotic nascent human beings the SCNT procedures that allowed Dolly’s birth – taking bioengineered cloned zygotic human beings and implanting them into a surrogate woman’s uterus for gestation and birth. Very recently there have been rare, and generally denounced, examples of births of human beings who began life as cloned zygotes implanted into the uterus of their mothers. The scientific motivation for these rare experiments is addressed later (See “Germline Genetic Engineering” on page 32).
Human reproductive cloning has not progressed to being an accepted strategy for assisted reproduction like in vitro fertilization (IVF). Cloned animals like Dolly have proven to develop disorders like premature aging that have not been observed for either animals or human beings conceived by IVF. The suspected cause of disorders in cloned animals is detrimental mutations in the DNA of the cloned cell nuclei used for their conception. When the DNA is replicated (copied) before cell division to make two new cells, errors can occur in the new versions of the genetic information. During the development and propagation of immortal cell lines, DNA copying errors (mutations) accumulate exponentially in the new cells produced. These mutations in the nuclei used for reproductive cloning can manifest as disorders in cloned individuals.
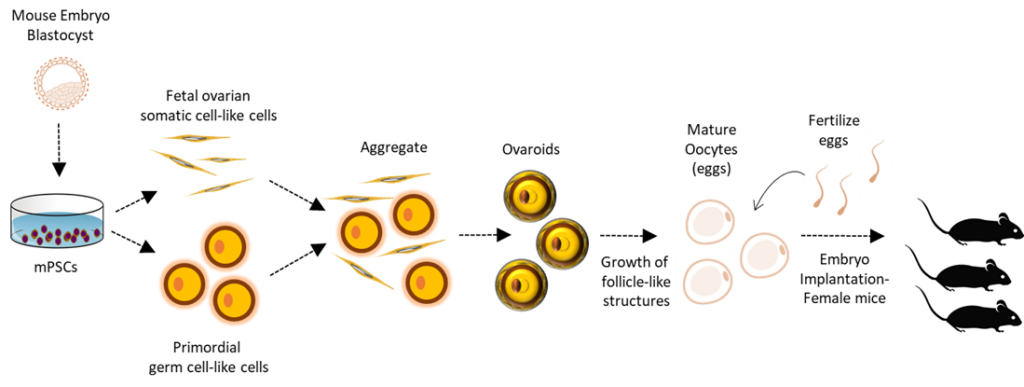
In Vitro Gametogenesis
IVG is an experimental technique for producing reproductive gamete cells in the laboratory. Recently, using pluripotent stem cells, researchers have been able to generate functional mouse ovary cells, which then produce female mouse eggs that can be fertilized successfully resulting in liveborn mice. This technique has not yet been done with humans. However, if at some point the technique is successful in humans, it could be used as a method for biomanufacturing a large number of human eggs for human cloning experiments. The supply of egg donors is currently a limitation in human cloning experiments. Producing bioengineered eggs would circumvent this problem.
Parthenogenesis
The developing female egg contains two maternal chromosomal copies (termed “diploid”), of which one maternal copy is normally eliminated during fertilization to make room for a paternal set of chromosomes from the male sperm. Parthenogenesis (in Greek meaning “virgin birth”) takes advantage of the diploid nature of the developing unfertilized egg. Parthenogenetic activation is a technique used to create living embryos (parthenotes) from unfertilized human eggs, without using sperm. Parthenogenesis uses chemicals to activate human development of the female egg in the absence of sperm fusion, while retaining both copies of maternal chromosomes. The resulting embryo contains two copies of each chromosome, both from the egg of the female donor. Unlike cloning, in which the conceived embryonic human being has the same genetic composition as the donor transferred nucleus (male and female chromosome copies), a parthenogenetic human being has two maternal copies of each chromosome (including two X chromosomes), thus creating female individuals only. Parthenogenetic embryonic human beings can have epigenetic and genetic defects, which can cause disorders and disease. Scientists are using parthenogenesis to create embryonic human beings for use in hESC research.
Experimentation Using Living Nascent Human Beings
There are many reasons given as justification by scientists who wish to pursue procurement and use of IVF human embryos and human cloning experimentation. These are human embryonic stem cell (hESC) research, therapeutic cloning, mitochondrial transfer, human embryo models, germline genetic engineering, and human-animal chimeras. Each is addressed in turn here. In each case, the experimentation is motivated by the attributes of human zygotes as nascent human beings, who will autonomously progress with their development from embryonic stages to fully mature human beings as long as supportive nutrients and conditions for growth are supplied.
Embryonic Stem Cell (hESC) Research
In the first hESC research, as described earlier (page 14), epiblasts were obtained from non-implanted embryonic nascent human beings conceived for IVF. A second source of embryonic nascent human beings who have been considered for hESC production is nascent human beings conceived by human cloning. Two early difficulties essentially precluded this type of experimentation with nascent human being subjects from progressing right at its start. First, early technical difficulties prevented scientists from achieving efficient conception of cloned human embryos; and second, at the same time, bioethics objections were raised against the cloning of nascent human beings for hESC research.
In efforts to address or circumvent bioethics objections to human cloning for therapeutic development (See next section), some scientists tried to devise strategies with alternative sources of eggs and/or sources of human nuclei with DNA genetically engineered to produce nascent human beings who would die soon after the stage at which epiblasts were removed to develop hESCs. Animal eggs (e.g., cow oocytes) proved incompatible with human DNA; and concerns arose that genetic mutations engineered to make nascent human beings inviable might also make the hESCs derived from them defective.
Research interest in hESCs has been motivated by their remarkable property of pluripotency. Human ESCs retain the ability of their epiblast precursors to produce many different cells of the embryonic and fetal body. In fact, hESCs are defined scientifically by their ability to produce cells that originate from all three of the germ layers that continue to the development of the organs and tissues in child human beings and adult human beings. The pluripotent development of ESCs can be restarted by switching them to culture conditions that allow embryoid body formation or by transplantation into experimental animals like mice.
The pluripotency of hESCs is commonly overstated. They are usually described as being “able to make any cell in the human body.” However, thus far, getting them to make mature cells found in child human beings and adult human beings has proved to be quite difficult. They have an intrinsic propensity to produce cells that are more like those found in embryonic and fetal organs and tissues. This shortcoming precluded their promised use to develop new medical therapies for child human beings and adult human beings.
A second problem preventing progress in developing new medical therapies with hESCs was recognized at the very beginning of permitting their use for human cell experimentation. They are intrinsically tumorigenic cells when they are not in the body of an embryonic nascent human being. Their tumorigenicity when outside of embryonic human beings is a manifestation of their pluripotency, which never evolved for function in the maturely developed organs and tissues of child human beings and adult human beings. The tumors formed are teratomas, which are chaotic masses of tissues caused by abnormal organizing of differentiated cells derived from all three germ layers of the human body.
In contrast to their behavior in foreign tissue environments, when ESCs are returned to an early embryonic environment they can fully regain their epiblast character and cause a conception to occur. A requisite embryonic environment has been established by an experimental technique called tetraploid complementation. In this procedure, after a zygotic embryo has undergone one cell division, producing two cells, it is given an electrical shock that induces the two cells to fuse and become one cell again. However, the new fused embryo has twice the full-complement of DNA. Because normal mammalian cells have two sets of DNA chromosomes, they are described as diploid cells. Since the new fused embryo has four sets of DNA chromosomes, it is termed tetraploid. Tetraploid embryos undergo a disrupted malformation of development. They complete development of the extraembryonic components of the fetus like the placenta, but a fetus does not develop.
When epiblasts are taken from an embryo to produce ESCs, the placental forming cells are left behind in the destroyed embryo. Therefore, alone, ESCs cannot complete formation of a viable fetus. However, scientists have shown for mouse and rat ESCs that if they are introduced into a tetraploid embryo, it will complement them and allow them to develop into a viable fetus. To date, tetraploid complementation with hESCs has not been pursued because of intense ethical and moral objection.
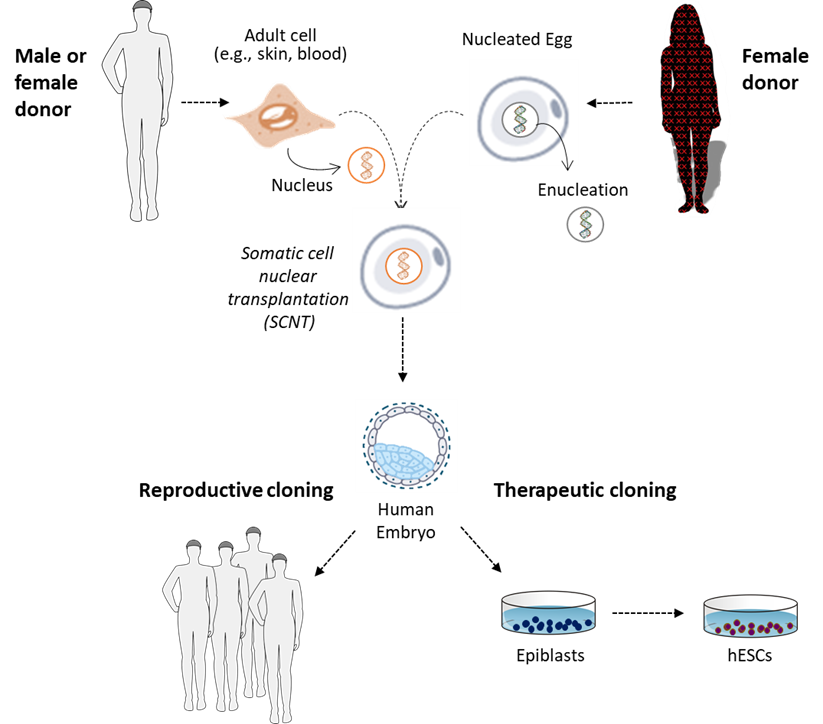
Therapeutic Cloning
As discussed earlier, by virtue to their origin from the inner cell mass of nascent human beings, hESC lines have unique properties that motivated interest in their use for research to develop new medical therapies. In the early days of human cloning research, it was considered as a possible new strategy for more rapid production of hESCs. Cloning for this purpose was called human therapeutic cloning, but it never gained significant adoption for research. Numerous factors were responsible for the failure of therapeutic cloning research. These included ethical objections to the cloning of nascent human beings in general; ethical and safety concerns about the required sourcing of human eggs from women; the teratoma-forming properties of hESCs, whether derived from IVF or cloned embryonic human beings; and objections that the same mutations that complicated reproductive cloning would also prevent normal function of therapeutically cloned tissues.
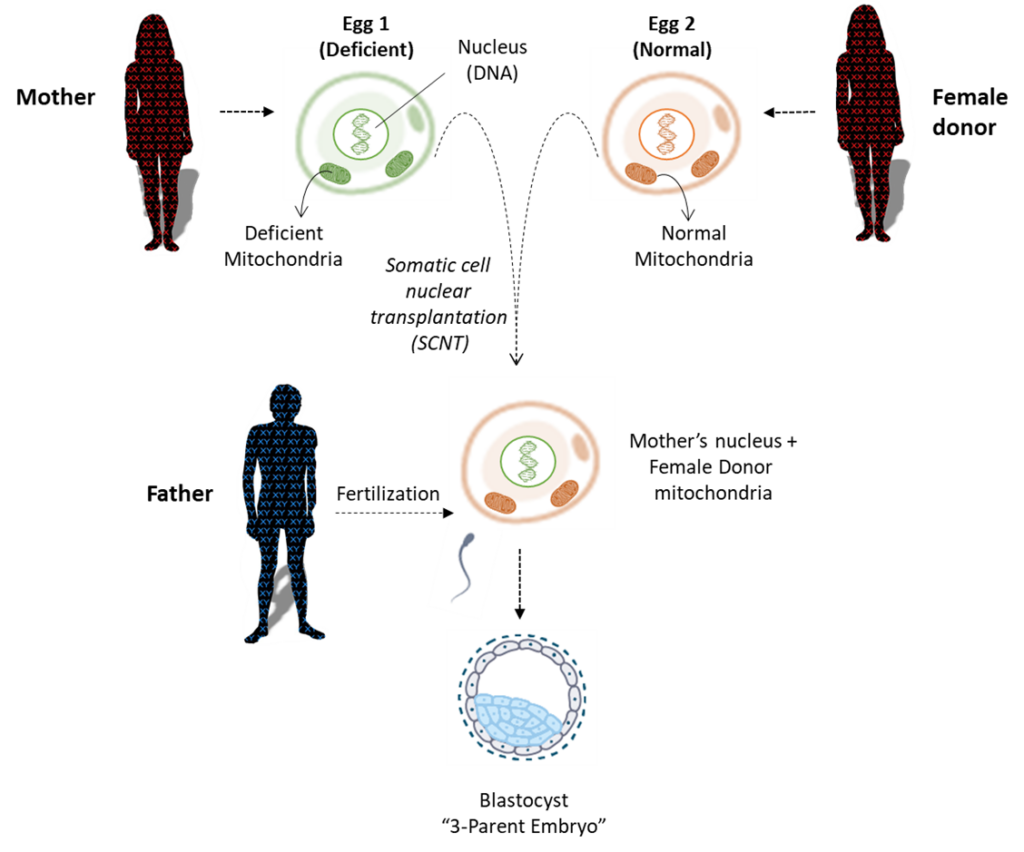
Mitochondrial Transfer (3-parent embryos)
Within each human cell are mitochondria organelles responsible for energy production (See earlier Human Cell figure on page 9). Mitochondria contain mitochondrial DNA, distinct from nuclear DNA, which is a genetic blueprint for a specific subset of cellular components found only in mitochondria. Like nuclear DNA, mitochondrial DNA is susceptible to mutations in the DNA code that can cause mitochondrial deficiency and diseases called mitochondrial disorders. Mitochondrial DNA has a unique pattern of inheritance. Because mitochondrial DNA only comes from the egg, it is inherited solely from the mother, whereas nuclear DNA is inherited from both mother and father.
The technique of mitochondrial transfer seeks to replace deficient mitochondria in a mother’s egg with normal mitochondria from a second woman donor’s egg. A fertilized egg that has undergone mitochondrial replacement will have nuclear DNA from the mother and father, plus mitochondrial DNA from a woman donor, hence the name “3-parent embryo”. Scientists are actively researching mitochondrial transfer techniques using donated embryos and gametes. The goal is to develop therapies that may increase fertility rates and reduce mitochondrial disease. This technique was reportedly successful in 2016, with the birth of the first “3-parent embryo” child in Mexico.
There are several different approaches used to complete the mitochondrial transfer and avoid deficient, disease-causing mitochondria that are described in more detail here. A simplified illustration of one method called “Maternal Spindle Transfer” is shown here.
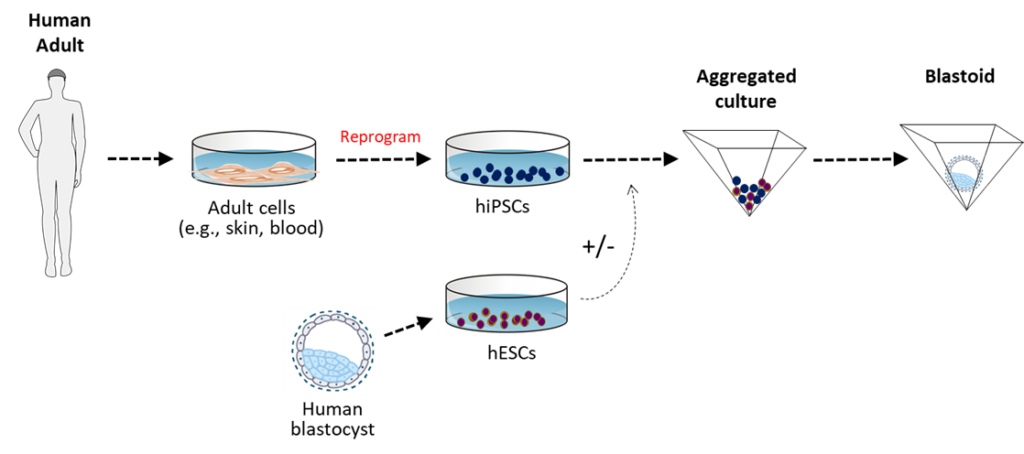
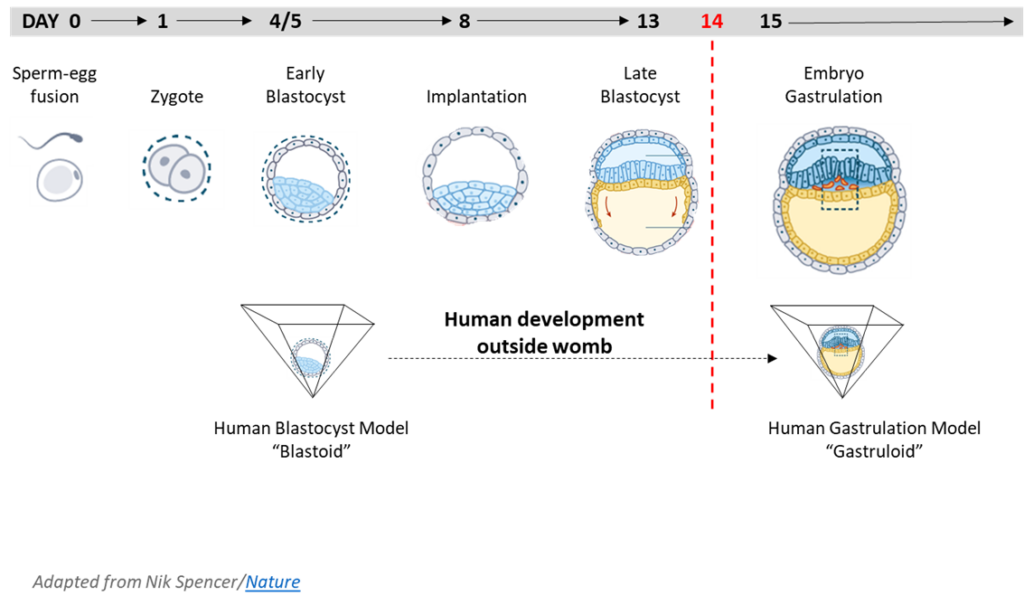
Human Embryo Models (Blastoids)
In 2021, earlier developments in mouse embryo research were applied by scientists to produce cloned embryonic human beings. Using hESC and hiPSC lines, described earlier, it is now possible to conceive nascent embryonic human beings who skip their zygotic age and begin life at the developmental age of blastula. Since these human beings are conceived from clonal cell lines, they are clones, being genetically identical. These embryonic human beings conceived from clonal hESC and hiPSC lines are now referred to as “induced blastoids (iBlastoids)” and “blastocyst-like structures.” When allowed to grow past the blastula stage, a more advanced “gastruloid” is produced corresponding to embryonic nascent human beings of gastrula age.
Growing Human Embryo Models Beyond 14 Days
The 14-day Rule was recently lifted in some countries (e.g., the UK and Australia), allowing scientists to culture human embryos beyond formation of the primitive streak, the equivalent of growing human beings into the 2nd week post-fertilization (or 4th week gestation) in pregnancy and beyond.
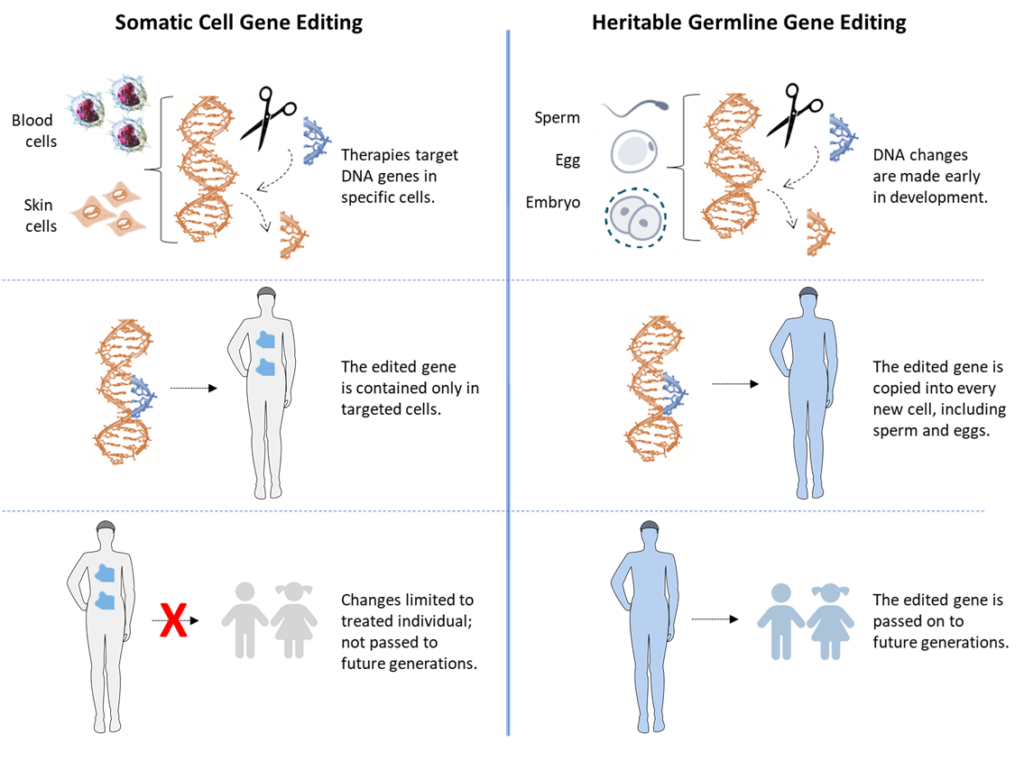
Genetic Engineering
A recent development in gene engineering called CRISPR (pronounced ‘kris-per) makes it possible for scientists to more efficiently and precisely make stable changes in the DNA code of cells. Changes in the DNA code made by CRISPR are called gene edits. Gene edits made in a cell’s DNA can result in changes in the properties and functions of the cell. Since a gene edit is inherited by all the descendants of a first edited cell, the descendant cells will also have the new properties and functions due to the gene edit.
There is much interest in using CRISPR technology to correct gene mutations that cause genetic diseases like beta thalassemia and sickle cell anemia. There are two strategies possible for this purpose. The first strategy is called somatic cell genetic engineering. In this case, CRISPR is used to correct inherited mutations in cells of the bodies of children and adults that cause an important deficiency, like being unable to make normal red blood cells. Blood-forming cells containing an inherited mutation that makes them defective are taken from the body, their gene mutation edited back to a good version, and then returned to the patient. This approach is currently being investigated in clinical trials for several genetic illness. An important requirement of such therapies is that the engineered cells are tissue stem cells. Tissue stem cells are the only cells of the body that are able to continuously renew the engineered therapeutic cells for a patient’s lifetime.
The second strategy is called heritable germline genetic engineering. In this case, CRISPR would be used to correct genetic mutations in human beings of zygotic age. Gene edits made in human beings at this stage of their development would be inherited by all of their mature cells when they grew up to be children and adults, including their gamete-forming reproductive cells. Therefore, although germline genetic engineering might be used to prevent inherited genetic diseases, it would also open the door to the birth of scientist-engineered human beings, whose modified genetic code, and the properties and functions it bestowed, could be passed on to their children. This aspect of this human experimentation is the basis for calling it germline. In addition to gene edits for disease intervention, germline genetic engineering would enable experiments with gene edits aimed to alter specific personal attributes, like height, eye color, musculature…and even intelligence. Germline editing was used by Chinese scientists to alter the genomes of twin baby girls born in 2018. The first germline-engineered human beings, these children were conceived with the intent to make their cells resistant to HIV infection in a way that will pass the trait on to future generations. The scientists ignored the risk that their genetic manipulation could also cause detrimental effects in these children and/or their future family generations.
A simplified illustration is shown here, comparing somatic cell gene editing to heritable germline gene editing.
Human-Animal Chimeric Embryo Research
Scientists are also attempting to create human-animal chimeras. The technique involves injecting human pluripotent stem cells from a human embryo (hESCs) or reprogrammed somatic cells (iPSCs) into animal blastocysts conceived by in vitro fertilization (IVF). In some strategies, the animal IVF embryo has been genetically-engineered to be unable to produce the animal organ that corresponds to the human organ of interest for production. Once human pluripotent cells fuse with the developing animal embryo, human cells begin growing with the animal (e.g., mouse, pig, monkey). At this time, only mouse-mouse chimeras have been successfully gestated to birth. No human-animal chimeras have been successfully gestated and born. However, various attempts have been tried as early “proof-of concept” studies, generating early pre-implantation human-animal embryo chimeras. The goal of this technique is to generate human-animal chimeras as a means of producing an unlimited supply of organs for human transplantation medicine needs.
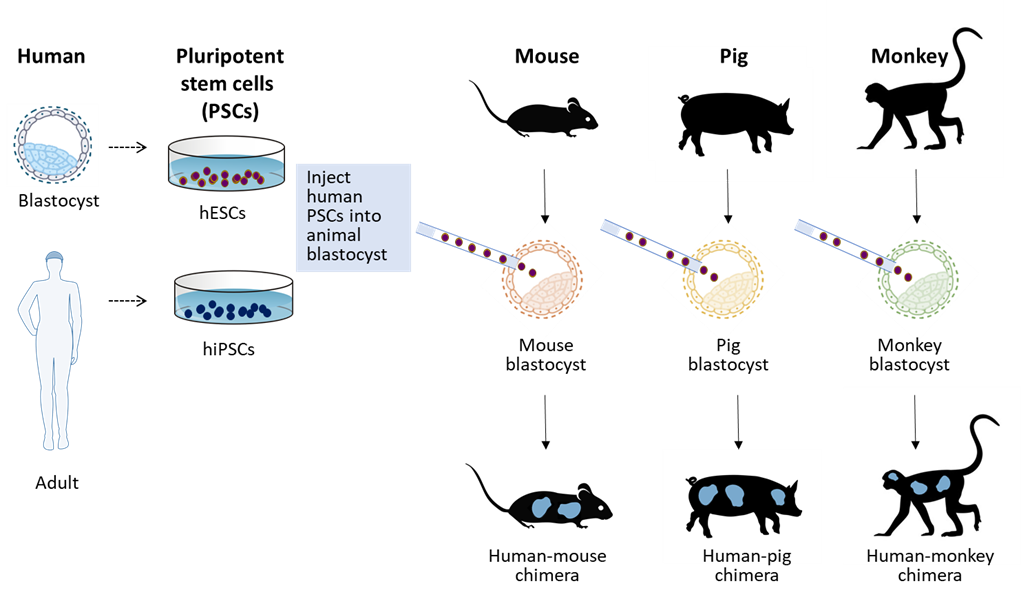
Table 1. State of the Ethical and Moral Permissibility of Human Experimentation Research